Нервная ткань
С нервной ткани (textus nervosus) построена центральная нервная система (головной и спинной мозг) и периферийная нервная система - нервы, нервные волокна с их конечными аппаратами, нервные узлы (ганглии). Нервная ткань состоит из нервных клеток - нейронов (нейроцитов) с особым строением и функцией и клеток нейроглии, выполняющих опорную, трофическую, защитную и разграничительную функции.
Нейроцитов, или нейрон (neurocytus, neuronum) является структурно-функциональной единицей нервной системы. Основными функциями нейрона являются: восприятие раздражения, анализ и трансформация этой информации в нервный (электрический) импульс или химический сигнал; передача и хранение этой информации, способность продуцировать биологически активные вещества. Благодаря таким функциям нейронов нервная ткань обеспечивает регуляцию и согласованную работу органов и систем организма, его адаптацию к условиям внутренней и внешней среды. Нейрон состоит из тела (перикариона), где информация обрабатывается, и отростков, отходящих от тела. Отростки является характерным структурным признаком нейронов, они обеспечивают проведение нервного импульса. Отростки есть двух видов - аксоны и дендриты. Аксон или нейрит (от греческого axis - ось), - это один длинный отросток длиной до 1,5 м, он заканчивается терминальным разветвлением. Этот отросток проводит нервный импульс от тела нейрона. Дендриты (от греческого dendron - дерево) - это короткие и многочисленные отростки, древовидные галузяться. Эти отростки проводят нервный импульс от окончаний к телу нейрона. Нервные клетки динамично поляризованные, то есть способны пропускать нервный импульс только в направлении от дендрита к аксона (рис. 1).
В зависимости от количества отростков нервные клетки делятся на: униполярные, имеющие только один отросток - аксон; биполярные, имеющих два отростки: аксон и дендрит; псевдоуниполярни нейроны имеют два отростки аксон и дендрит, но возле тела клетки эти два отростки настолько плотно прилегают друг к другу, что создается эффект одного отростка, но на определенном расстоянии эти отростки Т-образно расходятся. По функцией это чувствительные нейроны, в основном они расположены в чувствительных узлах спинномозговых и черепных нервов. Мультиполярные нейроны имеют многочисленные дендриты и один аксон, они преобладают в нервной ткани.
Размеры тела нервных клеток колеблются в пределах от 4-5 мкм до 130-140 мкм, а длина отростков колеблется от нескольких микрометров до 1 м и более. Форма тел нейронов, их размеры, число дендритов и степень их разветвления очень меняющимися в зависимости от локализации нейронов и выполняемой ими функции. Например, псевдоуниполярни нейроны имеют круглое тело, форма тел мультиполярные нейронов спинного мозга неправильная. Тела крупных пирамидных нейронов коры большого мозга имеют треугольную форму, а от них уходит много коротких дендритов. Аксон отходит от основания клетки. В отличие от дендритов, диаметр аксона не меняется. Грушевидные нейроны коры мозжечка имеют два крупных дендриты, интенсивно галузяться, а длинный аксон отходит от верхушки клетки.
Различают два типа мультиполярные нейронов: мультиполярные нейрон с длинным аксонов и большим количеством дендритов (клетка Гольджи 1-го типа) и мультиполярные нейрон с коротким аксоны, дендриты которого галузяться (клетки Гольджи II-го типа). Клетки 1-го типа расположены в симпатичных и парасимпатических нервных узлах. К нейронов 1-го типа также принадлежат крупные пирамидные нейроны коры большого мозга грушевидные нейроны коры мозжечка, двигательные нейроны спинного мозга. Эти нейроны передают нервные импульсы на большие расстояния. К нейронов II-го типа относятся клетки центральной нервной системы, передающие нервные импульсы соседним нейронам.
В сером веществе полушарий большого мозга и мозжечка нейроны располагаются слоями, а в других отделах нервной системы нервные клетки образуют скопления - ядра.
Нейроны - преимущественно одноядерные клетки. Два и больше ядер имеют некоторые нейроны, расположенные в нервных узлах автономной (вегетативной) нервной системы. Сферическое ядро диаметром около 17 мкм в большинстве нейронов занимает центральное положение (рис. 2). Гетерохроматин располагается равномерно по всему ядру, хорошо заметное базофильные ядрышко, в нейроплазми перикариону расположены многочисленные сферические или удлиненные митохондрии диаметром примерно 0,1 мкм. Часто в зоне комплекса Гольджи оказываются мультивезикулярни тельца.
Основными структурными признаками нейронов является наличие в нейроплазми многочисленных специальных органелл - нейрофибрил и скоплений хроматофильнои субстанции (вещество Ниссля, тигроид), состоящая из групп параллельных цистерн гранулярных эндоплазматической сетки и полирибосом, содержащие много РНК. Элементов агранулярный эндоплазматической сети в теле нейронов мало. Они есть только в аксонов и дендритах в виде трубочек, цистерн и пузырьков. Хроматофильна субстанция и свободные рибосомы располагаются по всей цитоплазме клетки и в дендритах, но она отсутствует в самом аксонов и его пригорке.
Между элементами эндоплазматической сети расположены многочисленные митохондрии, лизосомы, гранулы липофусцина. Митохондрии есть и в отростках нейрона. Центриолей в нейронах нет. Наружная поверхность цитолемы нейрона покрыта многочисленными синапсами и отростками астроцитов. Нейрофибрилы, переходящие в отростки, состоящие из микротрубочек диаметром около 20 нм и нейрофиламенты толщиной 7-10 нм. Нейрофибрилы формируют у перикариони густую трехмерную сетку, в которой расположены лизосомы и другие структуры. Нейрофибрилы обеспечивают прочность перикариону и отростков, осуществляют химическую интеграцию клетки.
РНК, синтезируемых в перикариони, транспортируются в отдаленные участки отростков. С помощью постоянного медленного транспорта макромолекул со скоростью 1-3 мм за сутки доставку делятся ферменты, принимающие участие в синтезе медиаторов в пресинаптические части синапсов, и белки цитоскелета. Быстрым антероградная транспортом поставляются пузырьки в синаптическую окончания со скоростью 400 мм за сутки Кроме того, существует ретроградный транспорт от окончаний аксона к перикари-она со скоростью 200-300 мм за сутки, с помощью которого крупные пузырьки переносят обломки структур и веществ, переваривают лизосомы. В дендритах проходит как медленное, так и быстрый транспорт.
В нейроплазми вдоль дендритов расположено много нейротрубочок, вытянутых митохондрий, а также есть небольшое количество цистерн агранулярный эндоплазматической сетки и нейрофиламенты. Вещество Ниссля есть и в крупных дендритах. Конечные отделы дендритов часто колбоподибно расширены.
Диаметр аксонов разных клеток (вместе с оболочками) колеблется в широких пределах от 1 до 20 мкм, но в одном отростке он всегда одинаков. Толстые аксоны проводят нервные импульсы быстрее, чем тонкие. Аксоны отходят от конического аксонного холма, вблизи которого от аксона ответвляются боковые ветки. Аксон заканчивается телодендроном - конечными разветвленными, образующих синапсы. Поверхность аксолемы (цитолемы) гладкая. Аксолема начального сегмента аксона и в области перетяжки Ранвье утолщенная. В аксоплазми являются тонкие удлиненные митохондрии, большое количество нейротрубочок и нейрофиламенты, пузырьки и трубочки агранулярный эндоплазматической сети, одинокие мультивезикулярни тельца. Рибосомы и элементы гранулярных эндоплазматической сети отсутствуют в аксоплазми, а есть только в цитоплазме бугорка аксона, где расположены пучки микротрубочек и немного нейрофиламенты.
Следовательно, нейроны воспринимают, проводят и передают электрические сигналы. Передача электрических сигналов обусловлена изменением мембранного потенциала, возникающее при перемещении через клеточную мембрану ионов натрия и калия благодаря функционированию натрий-калиевого насоса.
Нейроны, передающие информацию от места восприятия раздражение в центральную нервную систему, а затем до рабочего органа, связанные между собой с помощью многочисленных межклеточных контактов - синапсов (от греческого synapsis - связь), которые обеспечивают передачу нервного импульса от одного нейрона к другому. В синапсах происходит преобразование электрических сигналов в химические, а затем - химических сигналов в электрические. Нервный импульс вызывает, например, в парасимпатической окончании высвобождения посредника - нейромедиатора, который связывается с рецепторами постсинаптического полюса, и приводит к изменению его потенциала.
В зависимости от того, какие части нейронов спо-лучаються между собой, различают синапсы: Аксой-соматические, когда окончание аксона одного нейрона образует контакт с телом другого, Аксой-дендритные, когда аксоны вступают в контакт с дендриты, а также Аксой-аксонни, когда контактируют одноименные отростки. Такой синаптических устройство цепочек нейронов создает возможность для передачи информации в различные участки тела. При этом передача импульса осуществляется с помощью биологически активных веществ (химическая передача), а сами вещества, осуществляющие передачу, называются нейромедиаторами (от латинского mediator - посредник). Роль медиаторов выполняют две группы веществ: норадреналин, ацетилхолин, некоторые моноаминов (адреналин, серотонин, дофамин и аминокислоты - глицин, глутаминовая кислота) и нейропептиды (энкефалины, нейротензин, ангиотензин II, вазоактивный кишечный пептид, соматостатин, вещество II и т.п.). По функцией различают возбуждающие и тормозные синапсы.
В синапсе выделяют пресинаптические и постсинаптические части, которые разделены синаптической щели (рис. 3). Нервный импульс поступает по нервном окончанием в булавовидные пресинаптические часть, которая ограничена пресинаптических мембраной.
Цитоплазма пресинаптических части содержит большое количество круглых мембранных синаптических пузырьков диаметром от 4 до 20 нм с нейромедиатором. Когда нервный импульс достигает пресинаптических части, открываются кальциевые каналы. Ионы кальция проникают в цитоплазму пресинаптических части, их концентрация кратковременно возрастает. При увеличении содержания кальция синаптическую пузырьки, содержащие нейромедиатор, проникают в нейролему и нейромедиатор выделяется в синаптическую щель. Чем больше содержание ионов кальция, тем больше синаптических пузырьков выделяют нейромедиаторы. Постсинаптичниы потенциал возникает тогда, когда нейромедиатор связывается с рецепторами постсинаптические мембраны, а ее потенциал меняется. Таким образом, постсинаптические мембрана превращает химический стимул в электрический сигнал. Открываются Ка + - каналы и К + - каналы: ионы натрия поступают в постсинаптичниы полюс, а ионы калия выходят в синаптическую щель, в результате чего происходит деполяризация постсинаптические мембраны. Это приводит к изменению мембранного потенциала и возникновения электрического сигнала, величина которого прямо пропорциональна количеству нейромедиатора. Как только прекращается выделение нейромедиатора, пресинаптические окончания поглощает медиатор из синаптической щели. После этого рецепторы постсинаптические мембраны блокируются антагонистом и возвращаются в исходное состояние.
Нейроглии. Кроме нейронов, в нервной системе есть клетки нейроглии (neuroglia), выполняющих: опорную, трофическую, защитную, изолирующую, секреторную функции (рис. 4). Различают две группы нейроглии: глиоциты или макроглию (епендимоциты, астроциты и олигодендроциты), и микроглии.
Макроглия. Епендимоциты (ependymocytus) имеют кубическую или призматическую форму и одним слоем Выкладываем изнутри желудочки мозга и спинно-мозговой канал. Епендимоциты соединены между собой замыкающим (плотными) контактами и ленточными десмосомамы. От базальной поверхности некоторых епендимоцитив (таницитив) отходит отросток, проходящей между другими клетками, разветвляется и контактирует с базальной мембраной. Под слоем епендимоцитив лежит слой недифференцированных глиоцитив. Епендимоциты принимают участие в транспортных и обменных процессах, выполняют опорную и разграничительную функции.
Астроциты (astrocytus) являются основными глиальными элементами центральной нервной системы. Различают протоплазматичные и волокнистые астроциты. Протоплазматичные астроциты имеют звездчатые форму на их телах имеются многочисленные короткие выпячивания, которые служат опорой для отростков нейронов, а между ними и плазмолемою астроциты есть щель шириной около 20 нм. Многочисленные отростки протоплазматичных астроцитов заканчиваются на нейронах и в капиллярах. Отростки астроцитов образуют сетку, в которой расположены нейроны. Отростки таких астроцитов расширяются на концах, образуя широкие ножки, контактируют между собой. Эти ножки со всех сторон окружают нейроны и кровеносные капилляры, покрывая примерно 80% их поверхности (периваскулярная глиальная пограничная мембрана (membrana limitans gliae perivascularis). He покрываются этой мембраной лишь участки синапсов. Глиальная мембрана, которая образована расширенными концами отростков астроцитов, изолирует нейроны, создавая для них специфическое микроокружение. Отростки, достигающие расширенными окончаниями поверхности мозга, соединяясь между собой щель контактами (Нексус), образуют на ней сплошную поверхностную глиальную пограничную мембрану На этой пограничной мембране расположена базальная мембрана, которая отделяет ее от мягкой мозговой оболочки.
Волокнистые астроциты преобладают в белом веществе центральной нервной системы. Диаметр этих клеток равен примерно 10 мкм, они имеют многочисленные (20-40) разветвленные отростки. Отростки расположены между нервными волокнами, некоторые из них достигают кровеносных капилляров.
Олигодендроциты (oligodendrocytus) - это малые клетки овоиднои формы (6-8 мкм) с великим, богатым хроматином ядром, окруженным тонкой полоской цитоплазмы, в которой есть относительно мало органелл. Олигодендроциты располагаются вблизи нейронов и их отростков. От тел олигодендроцитов отходят многочисленные короткие конусообразные и широкие плоские трапециевидные миелин создающее отростки. Эти отростки формируют миелиновых слой нервных волокон, спирально накручуючись на них. Олигодендроциты, которые образуют миелиновую оболочку нервных волокон периферической нервной системы, называются лемоцитамы, нейролемоцитамы, или клетками Шванна.
Клетки микроглии (microglia), или клетки Гортега, составляют примерно 5% от клеток глии в белом веществе и 18% в сером веществе головного и спинного мозга. Это маленькие удлиненные клетки. От тела клетки отходят отростки, которые образуют вторичные и третичные короткие разветвления. Некоторые клетки микроглии контактируют с капиллярами. Эти клетки относятся к макрофагов, но они способны синтезировать иммуноглобулины.
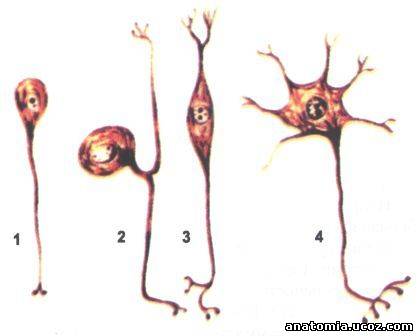
Рис. 1 Типы нейтроцитив
1 - униполярные; 2 - псевдоуниполярний; 3 - биполярный; 4 - мультиполярные.
1 - униполярные; 2 - псевдоуниполярний; 3 - биполярный; 4 - мультиполярные.
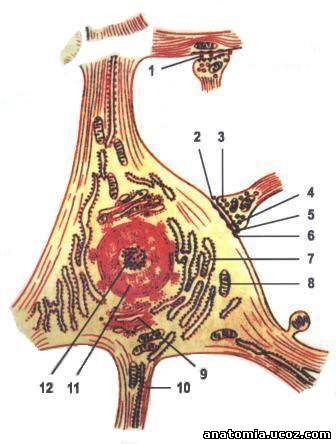
Рис. 2. Ультрамикроскопические строение нервной клетки.
1 - Аксой-дендритных синапс;
2 - Аксой-соматический синапс;
3 - пресинаптические пузырьки;
4 - пресинаптические мембрана;
5 - синаптическая щель;
6 - постсинаптические мембрана;
7 - эндоплазматической сети;
8 - митохондрия;
9 - комплекс Гольджи (пластинчатый комплекс); 10 - нейрофибрилы;
11 - ядро;
1 - Аксой-дендритных синапс;
2 - Аксой-соматический синапс;
3 - пресинаптические пузырьки;
4 - пресинаптические мембрана;
5 - синаптическая щель;
6 - постсинаптические мембрана;
7 - эндоплазматической сети;
8 - митохондрия;
9 - комплекс Гольджи (пластинчатый комплекс); 10 - нейрофибрилы;
11 - ядро;
12 - ядрышко.
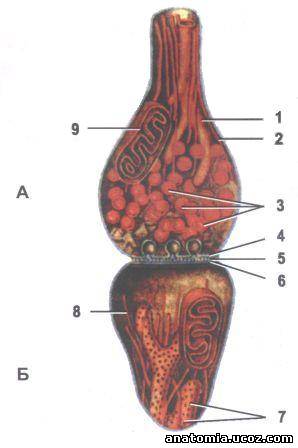
Рис. 3. Строение синапса.
А - пресинаптические часть;
Б - постсинаптические часть.
1 - агранулярный эндоплазматический сетка;
2 - нейротрубочка;
3 - синаптическую пузырьки;
4 - пресинаптических мембран с гексагональной сеткой;
5 - синаптическая щель;
6 - постсинаптические мембрана;
7 - гранулярная эндоплазматической сети;
8 - нейрофиламенты;
9 - митохондрия.
А - пресинаптические часть;
Б - постсинаптические часть.
1 - агранулярный эндоплазматический сетка;
2 - нейротрубочка;
3 - синаптическую пузырьки;
4 - пресинаптических мембран с гексагональной сеткой;
5 - синаптическая щель;
6 - постсинаптические мембрана;
7 - гранулярная эндоплазматической сети;
8 - нейрофиламенты;
9 - митохондрия.
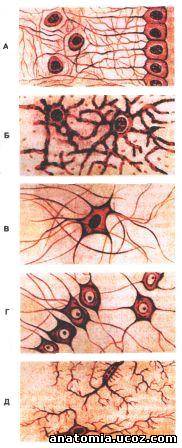
Рис. 4. Нейроглии
А - епендимоциты; Б - протоплазматични астроциты; В - волокнистые астроциты; Г - олигодендроциты; Д - микроглии.