Цитоплазма
Основными структурами цитоплазмы является гиалоплазма, органеллы и включения.
Гиалоплазма (от греческого hyalos - стекло), что занимает около 53-55% объема клетки, по физико-химическим параметрам является коллоидов. Гиалоплазма состоит из цитозоле (вода, ионы, органические молекулы) и цитоматриксу (сетка волокон диаметром 2-3 нм, состоящие из гликолипида, гликопротеинов и липопротеинов). В гиалоплазми содержатся органеллы и включения. В гиалоплазми осуществляется наиболее распространенный процесс выделения энергии - гликолиз, представляющий собой последовательную цепь ферментативных реакций, в результате которых шести углеводная молекула глюкозы расщепляется на два три углеводородные молекулы пировиноградной кислоты. При гликолитической расщеплении одной молекулы глюкозы образуется 4 молекулы АТФ.
Органеллы. Органеллами называют постоянные структуры цитоплазмы, имеющие характерную строение и выполняют специализированную функцию, необходимую для поддержания жизнедеятельности клетки. Они обеспечивают ее энергетический обмен, синтетические процессы, транспорт веществ и т.п.
Органеллы, которые есть во всех клетках, называются органеллами общего назначения, а органеллы, что есть только в специализированных клетках, - специальными. По наличию в составе органелл биологической мембраны различают мембранные и не мембранные органеллы.
Органеллы общего назначения. Мембранные органеллы. Каждая мембранная органелла является структурой цитоплазмы, которая ограничена биологической мембраной. К мембранных органелл принадлежат митохондрии, эндоплазматической сети, аппарат Гольджи (комплекс Гольджи, пластинчатый комплекс), лизосомы и Пероксисома.
Митохондрии обеспечивают процессы клеточного дыхания и превращают энергию, при этом образуется, в доступную форму для использования другими структурами клетки. Энергия накапливается в молекулах аденозинтрифосфорнои кислоты (АТФ), поэтому их образно называют "энергетическими станциями клетки".
Митохондрии, в отличие от других органелл, имеют собственную генетическую систему, которая необходима для самовоспроизведения и синтеза белков - ДНК, РНК и рибосом. Митохондрии размножаются в клетке путем бинарного деления. То есть, они являются органеллами, что само воспроизводятся. Но часть необходимых им белков кодируется ядерными генами и поступает в митохондрии с гиалоплазмы, поэтому митохондрии называют полуавтономных структурами.
Митохондрии имеют круглую, овальную или палочковидную форму, их длина составляет 0,3-5 мкм, а ширина - 0,2-1 мкм. Каждая митохондрия образована двумя биологическими мембранами - внешней и внутренней митохондриальной мембранами (рис.1). Между ними расположен между мембранный пространство шириной 10-20 нм. Внешняя мембрана ровная, внутренняя образует многочисленные складки - гребни (кристи). Благодаря гребня площадь внутренней мембраны значительно увеличивается.
Пространство, ограниченное внутренней митохондриальной мембраной, заполненный коллоидной митохондриальных матриксом. Он имеет мелкозернистую структуру и содержит многочисленные ферменты, обеспечивающие синтез АТФ. В матриксе помещен собственный генетический аппарат митохондрий, РНК и рибосомы. Гребни имеют вид складок или трубочек диаметром 20-60 нм (например, в клетках яичка, синтезирующих стероиды). Большинство гребней ориентированы поперек длинной оси митохондрий, но могут иметь и продольную ориентацию. Гребни (кристи) преимущественно не достигают противоположной стороны митохондриальной мембраны. Благодаря складкам площадь внутренней митохондриальной мембраны намного больше внешней. Так, поверхность одной митохондрии гепатоцита составляет примерно 13 мкм2, а площадь ее крист - около 16 мкм2. На внутренней поверхности гребней расположены многочисленные електроннощильни субмитохондриальни элементарные частицы (до 4 000 на 1 мк мембраны), имеющих форму гриба. Эти частицы содержат фермент АТФ-синтетазу, осуществляющий синтез и гидролиз АТФ.
В митохондриальном осмиофильному матриксе еще есть небольшие електроннощильни митохондриальную гранулы диаметром около 15 нм, состоящие из нерегулярных пластин (ламели), содержащих фосфолипиды, ионы кальция, магния, бария и стронция.
Количество, размеры и расположение митохондрий в клетке зависит от ее функции и потребности в энергии. Так, в одной печеночной клетке может быть до 2 500 митохондрий. Многочисленные крупные митохондрии есть в кардиомиоцитах и миосимпластах - поперечно исполосованная мышечных волокнах.
Эндоплазматическая сетка образует единую замкнутую внутренне жгутика циркуляционные систему, ограниченную мембраной толщиной примерно 60 нм, что образует многочисленные инвагинации и складки. На электронно фотографиях эндоплазматической сети имеет вид трубочек, плоских или круглых цистерн, мембранных пузырьков, сочетающиеся с перинуклеарним пространством клетки. На мембранах эндоплазматической сети происходит первичный синтез веществ, необходимых для жизнедеятельности клетки. их условно называют первичными, потому что молекулы этих веществ в дальнейшем используются в биохимических реакциях в других структурах.
Большинство веществ синтезируется на внешней поверхности мембраны эндоплазматической сетки. Потом эти вещества переносятся через мембрану внутрь этой сетки и по системе канальцев транспортируются к местам дальнейших биохимических превращений.
Есть два типа эндоплазматической сетки: гранулярная (шероховатая) эндоплазматической сети с диаметром каналец от 20 до 1 000 нм и агранулярный (гладкая) эндоплазматической сети с диаметром канальцев 50-100 нм.
К мембраны гранулярных эндоплазматической сети прикреплены многочисленные рибосомы. Они синтезируют белки, часть из которых используется самой клеткой, другие выводятся во внешнюю среду (например, синтез антител плазматическими клетками). Только гранулярная эндоплазматической сети образует биологические мембраны и ее элементы для всех органелл и других мембранных структур.
На поверхности агранулярный эндоплазматической сети нет рибосом. Между канальцами такой сетки часто расположены гранулы гликогена На мембранах эндо-плазматической сетки синтезируются углеводы и липиды, в частности гликоген и холестерин. Агранулярный эндоплазматический сетка обеспечивает выделение ионов хлора париетальных клетках эпителия желез желудка. Выполняя роль депо ионов кальция, эта сетка является необходимым звеном в механизме сокращения кардиомиоцитов и поперечнопосмугованих мышечных волокон. Чрезвычайно важную роль играют эти органеллы в детоксикации вредных для клетки химических соединений, например, гепатоциты обезвреживают токсичные вещества, поступающие из желудка и из кишечника по воротной вене в печень.
По канальцах эндоплазматической сети синтезированные вещества транспортируются в комплекс Гольджи.
Аппарат Гольджи (комплекс Гольджи, пластинчатый комплекс) представлен совокупностью цистерн, пузырьков, пластинок, трубочек и мешочков. При световой микроскопии он имеет вид сеточки. Чаще аппарат Гольджи состоит из трех мембранных элементов: уплощенным мешочков (цистерн), которые соединены между собой каналами, пузырьков, вакуолей. Концы цистерн расширены. От них отделяются пузырьки и вакуоли, окруженных мембраной и содержат различные вещества. Большинство мембранных пузырьков имеет диаметр 50-65 нм, а диаметр крупных вакуолей составляет 66-100 нм. Часть вакуолей содержит гидролитические ферменты и являются предшественниками лизосом.
Наиболее широкие уплощенные цистерны направлены в эндоплазматической сетки. В цистернах продолжается синтез полисахаридов, образуются комплексы белков, углеводов и липидов, которые транспортируются из одних цистерн в другие. В аппарате Гольджи завершается формирование продуктов синтетической деятельности клеток. Мембрана цистерн образует вырасти, куда перемещаются различные вещества. От выростов отделяются пузырьки, которые двигаются в разных направлениях в гиалоплазми. Часть пузырьков подходит к поверхности клетки и из них выводятся в межклеточное пространство синтезированные вещества.
Участок комплекса Гольджи, куда поступают вещества из эндоплазматической сети, называют цис-цистерной, противоположную - транс-цистерной. То есть комплекс Гольджи структурно и биохимический поляризованный. В направлении от цис-поверхности аппарата Гольджи к транс-поверхности увеличивается толщина мембран от 6 до 8 нм, а также содержание холестерина и углеводных компонентов в мембранных гликопротеин.
В процессе образования пузырьков расходуется значительное количество материала мембран, поэтому сбор биологических мембран является еще одной функцией комплекса Гольджи. Это сбор происходит из мембранных элементов, поступающих из эндоплазматической сети (рис. 2.).
Положение комплекса Гольджи в клетке обусловлено ее функциональной специализацией. В секреторных клетках эта органелла расположена между ядром и их апикальной поверхностью. В клетках эндокринных желез, секрет которых выводится в кровеносные капилляры, многочисленные элементы комплекса Гольджи расположены в периферийных отделах гиалоплазмы. В гепатоцитах (клетки паренхимы печени) структуры аппарата Гольджи располагаются группами: одни - у желчных путей, другие - около сосудов. В плазматических клетках (при световой микроскопии) пластинчатый комплекс занимает светлую зону возле ядра, окружен гранулярных эндоплазматической сети и на ее базофильными фоне выглядит как "светлое двор". Всегда вблизи комплекса Гольджи концентрируются митохондрии.
Лизосомы, которые отделяются от комплекса Гольджи, называют первичными. Каждая Лизосома является мембранным пузырьком диаметром 0,4-0,5 мкм, в котором содержится более 60 видов различных гидролитических ферментов в активированы состоянии (протеазы, липазы, фосфолипазы, Нуклеаза, гликозидазы, фосфатазы, в том числе кислая фосфатаза). Молекулы этих ферментов синтезируются на рибосомах гранулярных эндоплазматической сети, которые переносятся транспортными пузырьками в комплекс Гольджи.
В клетке лизосомы формируют лизосомных пространство, в котором поддерживается кислая среда (рН колеблется в пределах 3,5-5,0). Мембраны лизосом устойчивы к собственных ферментов и защищают цитоплазму клетки от их разрушительного воздействия. Эта функция обеспечивается особым расположением молекул в лизосомных мембране. При повреждениях лизосомных мембран нарушается ее проницаемость, что приводит к активации ферментов и деструктивных изменений клетки, вплоть до ее гибели.
Функция лизосом - внутриклеточный лизис (переваривание) высокомолекулярных соединений и частиц. Это могут быть собственные органеллы или включения, а также частицы, поступивших в клетку извне путем эндоцитоза. Такие частицы окружаются мембраной и называются фагосомамы.
Процесс внутриклеточного лизиса (переваривание) осуществляется в несколько этапов. Сначала первичная Лизосома сливается с фагосомою, и такой комплекс называют вторичной лизосомы (фаголизосомы). В вторичной лизосомы ферменты активируются и расщепляют полимеры, поступивших в клетку, до мономеров. Продукты расщепления транспортируются через лизосомную мембрану в цитозоле. Не переваренные вещества остаются в лизосомы и могут сохраняться в клетке очень долго в виде остаточного тельца. Остаточные тельца является включениями. Возможен и несколько иной путь преобразований: вещества фагосоми полностью расщепляются, после чего мембрана фагосомы распадается.
В процессе жизнедеятельности в клетках постоянно происходит перестройка структур, начиная от молекул и заканчивая органеллами. Вблизи поврежденных или таких, которые требуют замены, участков цитоплазмы, обычно возле комплекса Гольджи, образуется полумесячная двойная мембрана, которая растет и окружает поврежденные зоны. Затем эта структура сливается с лизосомами. В такой аутофагосоми происходит лизис захваченных структур.
Вторичные лизосомы могут объединяться между собой, а также с первичными лизосомами.Тогда возникают своеобразные вторичные лизосомы - мультивезикулярные тельца.
В некоторых случаях не переваренные остатки частиц накапливаются в лизосомах, что приводит к их "перегрузки". Тогда не переваренные остатки путем экзоцитоза выводятся в межклеточное пространство, а это может привести к деструктивным изменениям окружающих клеток.
Пероксисома (микротильця) являются мембранными пузырьками диаметром 0,2-0,5 мкм. Как и лизосомы, они отделяются от транс-цистерн комплекса Гольджи. Есть две формы пероксисом. Небольшие Пероксисома (диаметром 0,15-0,25 мкм) содержат мелкозернистый осмиофильний материал и морфологически почти не отличаются от первичных лизосом. Большие Пероксисома (диаметром более 0,25 мкм) есть только в клетках некоторых органов (печень, почка). Для них характерна кристалловидная сердцевина, созданной ферментами в концентрированном виде. У пероксисом могут располагаться и другие мембранные микротильця диаметром от 0,5 до 10 мкм, содержащих различные ферменты.
Пероксисома содержат ферменты пероксидазу, каталазу и оксидазу D-аминокислот. Пероксидаза участвует в обмене липидов, холестерина, пуринов, а также перекисных соединений, в частности перекиси водорода, который токсичен для клетки. Для биохимических реакций в Пероксисома используется молекулярный кислород. Пероксисома нейтрализуют много токсичных соединений, например, этанол.
Немембранни органеллы. К немембранных органелл принадлежат цитоскелет, клеточный центр и рибосомы.
Цитоскелет включает микротрубочки, микрофиламенты и промежуточные филаменты.
Микротрубочки пронизывают всю цитоплазму клетки. Каждая микротрубочки представляет собой полый цилиндр диаметром 20-30 нм. Стенка микротрубочки имеет толщиной 6-8 нм. Многие микротрубочек расположены радиально к центриолей. Микротрубочки образуют опорные структуры цитоскелета. Часть микротрубочек располагается согласно сил сжатия и натяжения, действующие на клетку. То есть такая конструкция цитоскелета соответствует принципам биомеханики. Особенно это заметно в клетках эпителиальных тканей, разграничивают различные среды организма. Микротрубочки участвуют в транспорте веществ внутри клетки. Структура стенки микротрубочек может изменяться в зависимости от факторов воздействия на клетку.
Промежуточные филаменты образованы длинными белковыми молекулами толщиной 8-10 нм. Они тоньше от микротрубочек, но толще микрофиламенты, поэтому и получили такое название.
Микрофиламенты - это белковые нити толщиной примерно 5 нм. Большинство микрофиламентов образована из молекул актиний, которых около 10 видов. Актиновые филаменты могут группироваться в пучки, которые образуют собственно опорные структуры цитоскелета. Филаменты актина способны также образовывать комплексы с полимерными молекулами белка миозина. Микрофиламенты при соединении с плазмолемою способны изменять ее конфигурацию. Это способствует поступлению веществ в клетку путем пиноцитоза и фагоцитоза.
Центросома (клеточный центр) (рис. 3) образовано из двух расположенных рядом центриолей (диплосома), которые окружены центросферою. Центросфера является гиаплазмою, которая не содержит органелл. Обе центриоли диплосомы расположенные между собой под углом. Основной функцией центросома является участие в образовании и росте новых микротрубочек. Перестройка структур микротрубочек происходит постоянно. Она также обеспечивает расхождение хромосом при делении клетки. В интерфазе клетки центросома размещена возле ядра. Эта органелла отсутствует в яйцеклетке.
Каждая центриоль представлена цилиндром, стенки которого состоят из 9 комплексов микротрубочек длиной 0,5 мкм и диаметром 0,25 мкм. Каждый комплекс образованный из 3 микротрубочек, поэтому называется триплета. Триплеты, расположенных между собой под углом приблизительно 50 °, состоят из 3 микротрубочек диаметром около 20 нм (изнутри наружу): полной А, неполных В и С. От трубочки А отходят две "ручки". Одна из них направлена в трубочки соседнего триплета другая - к центру цилиндра. Внутренние "ручки" напоминают фигуру звезды или спиц колеса. Центриоли - саморегулирующиеся структуры, удваиваются в процессе клеточного цикла. При митотического разделении обе центриоли расходятся к противоположным полюсам клетки. Центриоли участвуют в образовании митотического веретена, базальных телец ресничек и жгутиков.
Рибосомы - это тельца диаметром 20-30 нм, состоящие из двух субъединиц - большой и малой. Каждая субъединица представлена комплексом с рибосомальной РНК (рРНК) и белков. Основная функция рибосом - сборка белковых молекул из аминокислот, которые поставляют транспортные РНК (тРНК). Между субъединица рибосомы есть щель, в которой размещена молекула информационной РНК (иРНК), а на большой субъединицы - бороздка, в которой формируется новый белковый цепь. Последовательность сборки аминокислот в белковый цепочка соответствует нуклеотидами в цепи иРНК. Таким способом осуществляется трансляция генетической информации.
Рибосомы могут располагаться в гиалоплазми одиночно или группами в виде розеток, спиралей, завитков. Такие группы называют полирибосомамы (полисомамы). Значительная часть рибосом прикреплена к мембранам, к поверхности эндоплазматической сети и к внешней мембраны кариотекы. Свободные рибосомы синтезируют белок, необходимый для жизнедеятельности самой клетки, а рибосомы, прикрепленные к мембране - белок, который выводится из клетки. Количество рибосом в клетке может достигать десятков миллионов.
Среди специальных органелл выделяют мигающие реснички и жгутики. Подробное описание других специальных органелл приведены в соответствующих разделах по цитологии, эмбриологии, гистологии.
Реснички - это вырост плазмолемы клетки длиной 5-10 мкм и диаметром 200 им (рис. 4). В основе реснички на уровне кортикального слоя цитоплазмы расположенное базальное тельце (кинетосома), которое образовано девятью периферийными триплеты коротких микротрубочек, окружающих один центральный триплет. Над базальных тельцем в вийци расположена аксонема (осевой филамент), состоящий из девяти пар (дуплетов) периферийных микротрубочек, формирующих цилиндр диаметром около 150 нм. В центре цилиндра является центральная пара микротрубочек. На поперечном сечении реснички выглядит как колесо с 9 спицами.
Все реснички клетки делают координированные колебательные движения. их количество в клетке достигает нескольких сотен. Например, к 250 ресничек длиной 5-15 мкм и диаметром 0,15-0,25 мкм расположены на апикальной поверхности реснитчатым эпителиоцитов верхних дыхательных путей, маточных труб, семенных трубочек яичка. Движение ресничек помогает транспортировать жидкость или частицы по трубчатых структурах.
Жгутики по конструкции напоминают реснички, но их длина достигает 150 мкм, а диаметр составляет 200 нм. Изменяя свое положение, они также выполняют функцию движения. Этот процесс обеспечивается благодаря скольжению дуплетов микротрубочек между собой, что обусловлено изменениями конфигурации молекул белка динеину.
Включение. Включениями называют непостоянные структурные компоненты цитоплазмы клетки, возникают как продукты ее метаболизма или попадают в клетку извне. Среди включений условно различают: трофические, пигментные и секреторные. К трофических включений принадлежат капли жира, гранулы гликогена, белковые гранулы. Эти вещества накапливаются в клетке, а затем используются ею при определенных функциональных потребностях. Пигментные включения могут быть окружены мембраной.
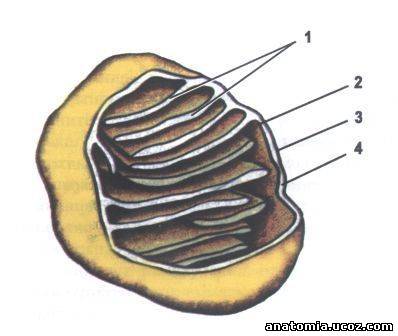
Рис. 1. Строение митохондрий.
- Кристи;
- Внутренняя митохондриальная мембрана;
- Внешняя митохондриальная мембрана;
- Межмембранную пространство
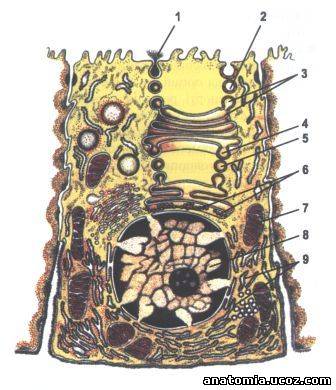
Рис. 2. Строение секреторного пути и восстановление мембран.
1 - экзоцитоз;
2 - формирование секреторных пузырьков;
3 - терминальное гликозирование;
4 - образование липидов;
5 - сульфатирование;
6 - синтез белка;
7 - митохондрия;
8 - ядро;
9 - гранулярная эндоплазматическая сеть.
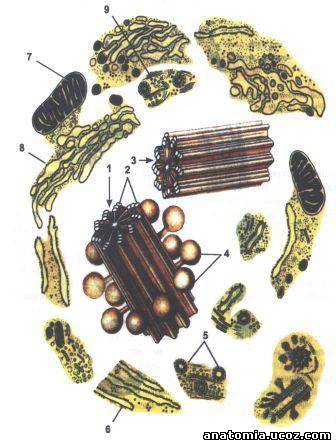
Рис. 3. Строение центросома (клеточного центра).
1 - центросфера;
2 - центриоль на поперечном разрезе (триплеты
микротрубочек, осевой компонент - структура "колеса телеги");
3 - центриоль;
4 - сателитарные тельца;
5 - окаймленные пузырьки;
6 - гранулярная эндоплазматической сети;
7 - митохондрия;
8 - комплекс Гольджи (пластинчатый комплекс);
9 - микротрубочки.
1 - центросфера;
2 - центриоль на поперечном разрезе (триплеты
микротрубочек, осевой компонент - структура "колеса телеги");
3 - центриоль;
4 - сателитарные тельца;
5 - окаймленные пузырьки;
6 - гранулярная эндоплазматической сети;
7 - митохондрия;
8 - комплекс Гольджи (пластинчатый комплекс);
9 - микротрубочки.
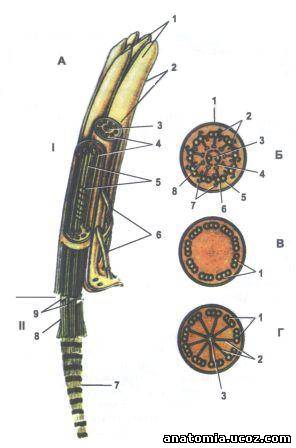
Рис.4. Строение реснички.
А - реснички на продольно-поперечном разрезе:
I - Зовнишньоклитинна часть;
II - Внутриклеточная часть:
1 - реснички;
2 - плазмолема;
3 - центральная пара микротрубочек (дуплет);
4 - периферийные пары микротрубочек (дуплеты);
5 - центральная влагалище;
6 - микроворсинки;
7 - базальный корешок;
8 - базальное тельце;
9 - внешние микротрубочки.
А - реснички на продольно-поперечном разрезе:
I - Зовнишньоклитинна часть;
II - Внутриклеточная часть:
1 - реснички;
2 - плазмолема;
3 - центральная пара микротрубочек (дуплет);
4 - периферийные пары микротрубочек (дуплеты);
5 - центральная влагалище;
6 - микроворсинки;
7 - базальный корешок;
8 - базальное тельце;
9 - внешние микротрубочки.
Б - поперечный разрез реснички через зовнишньоклитинну часть:
1 - плазмолема;
2 - периферийные пары микротрубочек (дуплеты);
3 - центральная пара микротрубочек (дуплет);
4 - центральная влагалище;
5 - субфибрила;
6 - субфибрила;
7 - динеинови «ручки»;
8 - радиальная спица.
В -поперечный разрез реснички через промежуточную участок, что соединяющий аксонему из базальных тельцем:
1 - триплеты микротрубочек.
Г - срез через базальное тельце реснички:
1 - триплет микротрубочек;
2 - радиальные спицы;
3 - центральное влагалище.