Нервные волокна (fibra newi, neurofibra) - это отростки нервных клеток, которые окружены оболочкой из олигодендроцитив (нейролемоцитив, или клеток Шванна). Различают миелиновые и безмиелиновые нервные волокна.
Безмиелиновые нервные волокна тонкие, их диаметр составляет 1-4 мкм. Они типичны для автономной (вегетативной) нервной системы. Характерной их структурным признаком является то, что отростки нейронов (осевые цилиндры) прогибает плазматической мембраной олигодендроцита (нейролемоцита) и погружаются в него, образуется глиальные муфта вокруг нервного отростка (рис. 1). В одну клетку Шванна может быть погружено много нервных волокон. Однако некоторые волокна не покрыты со всех сторон олигодендроцитамы. Группа безмиелинових нервных волокон, которая связана с одним нейролемоцитом, покрытая ендоневрием, образованным базальной мембраной нейролемоцита и тонкой сеточкой из коллагеновых и ретикулярных микрофибрилл.
Миелиновые нервные волокна имеют сложное строение потому нейролемоцит (клетка Шванна) спирально накручивается на осевой цилиндр (аксон) нервной клетки. При этом цитоплазма и ядро нейролемоцита оттеснены на периферию в поверхностные отделы оболочки (рис. 2). Это толстые волокна диаметром от 1 до 20 мкм. Они являются составной частью центральной и периферической нервной системы. Каждый нейролемоцит охватывает только часть осевого цилиндра длиной примерно 1 мм, образуя межузловых сегмент нервного волокна. Миелин - это многократно закрученный двойной слой цитоплазматической мембраны нейролемоцита. Толстая и плотная миелиновая оболочка, богатая липидами, изолирует нервное волокно и предотвращает "утечки" электрического тока (нервного импульса) с аксолемы. Внешняя оболочка осевого цилиндра образована цитолемою нейролемоцита, его базальной мембраной и тонкой сеточкой из ретикулярных и коллагеновых фибрилл. В темной миелиновых оболочке (при окраске препаратов осмиево кислотой) расположены узкие светлые косые линии - насечки миелина. Эти структуры связаны с процессом формирования миелиновой оболочки. Двойная складка плазмолемы нейролемоцита, что концентрически расположена вокруг осевого цилиндра, называется мезаксоном. Каждый завиток мезаксона имеет толщиной 8-12 нм. Насечки миелина соответствуют местам, где завитки мезаксона раздвинуты цитоплазмой лемоцитов. На границе между двумя соседними нейролемоцитамы образуется сужение нервного волокна - узловая перетяжка нервного волокна (перетяжка Ранвье), шириной в среднем 0,5 мкм, где миелиновая оболочка отсутствует (рис.3 ). Здесь аксолема контактирует с отростками, переплетающихся между собой, нейролемоцитамы и их базальной мембраной. В этом месте сконцентрировано большинство натриевых каналов (3000-5000 на 1 мкм), а в цитолеми, покрытой миелина, они практически отсутствуют. Междоузлия сегменты покрыты миелина, поэтому время проведения по ним нервного импульса приближается к нулю.
В аксолеми на уровнях перетяжек Ранвье генерируется нервный импульс, стремительно проводится в соседнюю перетяжки, в которой возбуждается следующий потенциал действия. Такой способ проведения импульса называется сальтаторним (скачкообразным). В миелиновых нервных волокнах скорость проведения нервного импульса высокая - от 5 до 120 м / с. Безмиелинови нервные волокна проводят нервный импульс со скоростью всего 1-2 м / с, потому что волна деполяризации движется по всей плазмолеми, не прерываясь.
Между нейронами и глиоцитамы является межклеточные щели шириной 15-20 нм. Они заполнены основной межклеточной веществом из мукополисахаридов и других соединений, обеспечивающих диффузию кислорода и питательных веществ. Все межклеточные щели соединяются между собой и образуют межклеточное (интерстициальное) пространство, на который приходится 17-20% общего объема мозга.
В зависимости от функции выделяют три основных типа нейронов: афферентные, ассоциативные и эфферентные. Афферентные нейроны (от латинского affevens - тот, что приносит) по функции чувствительны (рецепторными) клетками, они проводят нервные импульсы от органов и тканей в мозг. Тела таких нейронов сосредоточены в нервных узлах (ганглиях), расположенные вне центральной нервной системой. Как правило, это биполярные (псевдоуниполярные) нейроны. Долгое дендрит такой нервной клетки направляется на периферию и заканчивается соответствующим рецептором, который способен трансформировать энергию внешнего раздражителя в нервный импульс. Второй отросток (аксон) идет в головной или спинной мозг, принося соответствующую информацию.
Рецепторы (receptores) в зависимости от их локализации разделяют на: екстерорецепторы, воспринимающие внешние раздражения, - они расположены в наружных покровах тела (коже и слизистых оболочках, в органах чувств); интерорецепторы, которые разделяют на те, которые реагируют на изменение химического состава внутренней среды (хеморецепторы), давления в тканях и органах (барорецепторов, механорецепторов); проприорецепторы, расположенных в мышц, сухожилий, связках, фасциях, костях, суставных капсулах и т.д. и реагируют на структурно-функциональные изменения в опорно-двигательном аппарате .
В зависимости от характера восприятия раздражения различают: терморецепторы, механорецепторов, барорецепторов, хеморецепторы и ноцирецепторы. Терморецепторы воспринимают изменение температуры. Механорецепторов реагируют на различные виды механических воздействий (прикосновение к коже, ее сжатия и др.), ноцирецепторы воспринимают болевые раздражения. Барорецепторов воспринимают изменения давления, а хеморецепторы реагируют на воздействие химических факторов. Среди нервных окончаний различают свободные, состоящие только из разветвлений осевого цилиндра, и несвободные, или конечные нервные тельца, содержащие окончания нервного волокна и клетки глии.
Свободные нервные окончания (termindtio nervi liberae) расположены в коже. Подходя к эпидермиса, нервное волокно теряет миелин и проникает через базальную мембрану в эпителиальный слой. При этом базальные мембраны эпителия и нейролемоцитив переходят друг в друга. Нервные волокна разветвляются между эпителиоцитами, доходя до зернистого слоя; их веточки диаметром менее 0,2 мкм колбоподибно расширяются на концах. Аналогичные конечные нервные окончания есть в эпителии слизистых оболочек и роговице глаза. Конечные свободные нервные окончания воспринимают боль, тепло и холод.
В эпидермисе свободные нервные окончания подходят к изменившимся эпителиоцитов (клеток Меркеля), которые имеют много пальцеподибних выростов. В цитоплазме этих клеток много електроннощильних мембранных гранул диаметром приблизительно 100 нм. Нервное окончание расширяется и образует из клеток Меркеля синапсоподибний контакт. Эти окончания является механорецепторов и воспринимают давление.
Барорецепторов - это разветвленные свободные нервные окончания, расположенные в адвентицийному слое стенки крупных артерий грудной полости и шеи. Важнейшие рецепторы залегают в стенках дуги аорты и сонной пазухи. При растяжении стенки артерии под влиянием артериального давления, что меняется, они возбуждаются, а закодированная информация о величине давления крови передается в центральную нервную систему. Таким образом, барорецепторов обеспечивают регуляцию артериального давления по принципу обратной связи.
Несвободные нервные окончания могут быть капсулированных, т.е. окружены соединительнотканной капсулой, и некапсульованимы, которые не имеют капсулы. Некапсульовани нервные окончания (corpusculum nervosum попcapsulatum) расположены в соединительной ткани. К ним относятся также окончание в волосяных фолликулах. Капсулированных нервными окончаниями (corpusculum nervosum capsulatum) является осязательные тельца (осязательные тельца Мейснера), пластинчатые тельца (тельца Фатер-Пачино), цибулиноподибни тельца (тельца Гольджи-Маццони). Все эти нервные окончания есть механорецепторов. В группу капсулированных нервных окончаний принадлежат колбочки Краузе, которые являются терморецепторы.
Каждый волосяной фолликул окружен расширенными несвободными нервными окончаниями, которые расположены снаружи от соединительнотканных элементов влагалища корня волоса. Базальная мембрана нервных окончаний переходит в базальную мембрану внешней эпителиальной влагалища. Уплощенное нервное окончание залегает между двумя клетками Шванна. При движении во-лосины возникает нервный импульс и передается в центральную нервную систему.
Пластинчатые тельца (тельца Фатер-Пачино) - крупнейшие из всех капсулированных нервных окончаний. Они имеют овальную форму, длину до 2 мм и толщину 0,5 мм. Расположенные в соединительной ткани внутренних органов и в подкожной основе (рис. 4,5). Тельце извне покрыто соединительнотканной капсулой, имеющей пластинчатые строение. Во соединительно тканевой оболочкой размещена внешняя луковица, состоящая из 10-60 концентрических пластинок, образованных уплощенным периневральным эпителиоидных клетками, соединенными между собой десмосомамы и замыкающими зонами.
Пространства между пластинками заполнены жидкостью, коллагеновыми микрофибрилл и редкими кровеносными капиллярами. Войдя в тельце, нервное волокно теряет миелиновую оболочку, но внутри тельца оно окружено клетками Шванна, формирующие внутреннюю луковицу. Нервное волокно, колбоподибно заканчивается, содержит много маленьких сферических митохондрий и светлых синаптических пузырьков. При сжатии тельца в нервном окончании возникает нервный импульс.
Осязательные тельца (осязательные тельца Мейснера) длина 50-160 мкм, шириной примерно бОмкм, имеют овальную или цилиндрическую форму, их много в сосочковыми слое кожи пальцев рук и стопы. Тельце образованное многочисленными удлиненными или грушевидными клетками Шванна, лежащие одна на одной. Нервное волокно, входя в тельце, теряет миелин и располагается между цитоплазматическими отростками. Каждое волокно заканчивается колбоподибним расширением. Периневрий переходит в капсулу, которая образована несколькими слоями эпителиоидных периневральным клеток. Сжатие этих клеток передается осязательную тельцу, вызывая деформацию его клеток, что приводит к возникновению импульса в нервных волокнах. То есть, осязательные тельца Мейснера является механорецепторов тактильной чувствительности.
Цибулиноподибни тельца (колбочки Краузе) расположены в коже, коньюнктивы глаза, слизистой оболочке полости рта. Сферические тельца окружены тонкой соединительнотканной капсулой, содержащей много коллагеновых волокон и фибробластов. Войдя в капсулу, нервное волокно теряет миелиновую оболочку и разветвляется в центре колбы на многочисленные веточки. Колбочки Краузе воспринимают холод, возможно, они есть и механорецепторов.
Проприорецепторы реагируют на напряжение мышц, сухожилий и суставных капсул, возникающие при движениях и изменении положения тела в пространстве. Это нервно-мышечные и нервно-сухожилий веретена, расположенные в брюшке мышц или в их сухожилия.
Большое (длиной 3-5 мм и толщиной до 0,5 мм) нервно-мышечное веретено окружено плотной соединительнотканной внутренней капсулой и состоит из 10-12 поперечно исполосованная мышечных волокон, а также нервных волокон. Наружная капсула веретена формируется из поперечно исполосованная мышечных волокон. Продольно расположенные веретена растягиваются вместе с мышцей и передают информацию о длине и степень растяжения мышцы.
Вставные (ассоциативные или кондукторни) нейроны передают нервный импульс от чувствительного (центростремительной) нейрона к двигательного или секреторного (центробежного) нейрона.
Тела эфферентных (эффекторных; двигательных; секреторных) нейронов (от латинского effevens - тот, который выносит) расположены в центральной нервной системе, симпатичных и парасимпатических нервных узлах. Аксоны эфферентных нейронов проводят нервные импульсы в рабочие органы (мышц, желез), а также ко всем органам и тканям, регулируя таким образом обмен веществ. Выделяют также двигательные и секреторные окончания аксонов эфферентных нейронов. Аксоны двигательных нейронов разветвляются, каждая такая ветка иннервируют определенное количество мышечных волокон. Двигательные (моторные) нервные окончания располагаются на мышечных волокнах. Окончание одного двигательного нейрона и поперечно исполосованная мышечное волокно, что им иннервуеться, образуют двигательную единицу - мион.
Каждое мышечное волокно иннервируют веточка аксона двигательного нейрона, которая заканчивается на поверхности волокна, образуя нервно-мышечное окончание, аборухову конечную пластинку - моторную бляшку (рис. 6). Веточки аксонов, контактирующие с мышечным волок-ном, уже не имеют миелиновой оболочки. Каждое такое нервное окончание прогибает сарколему мышечного волокна и образует нервно-мышечный синапс. Нервно-мышечный синапс состоит из пресинаптических мембраны (аксолема) и постсинаптические мембраны (сарколема мышечного волокна), разделенных синаптической щели (рис. 7).
Нервный импульс, достигающий нервно-мышечного синапса, вызывает выход в синаптическую щель ацетилхолина, который связывается со специфическими рецепторами мембраны мышечного волокна, что приводит к открытию натриевого и калиевого каналов. Это вызывает местную деполяризацию постсинаптические мембраны, а электрический потенциал запускает механизм сокращения мышечного волокна.
Нервные окончания в гладкой мышечной ткани имеют более простую строение, образуя на поверхности миоциты характерные расширения, в которых есть синаптическую пузырьки с норадреналин и дофамин. Большинство окончаний аксонов контактируют с базальной мембраной гладкого миоциты, лишь незначительное количество нервных окончаний пронизывает базальную мембрану Тогда аксолема отделена от плазмолемы миоциты щелью шириной около 10 нм.
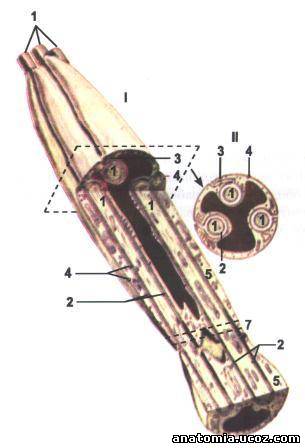
Рис. 1. Строение безмиелинового нервного волокна
I - продольный разрез;
II - поперечный разрез.
1 - осевые цилиндры;
2 - аксолема;
3 - мезаксон;
4 - клеточная оболочка лемоцита (нейролемоцита; клетки Шванна);
5 - цитоплазма лемоцита;
6 - ядро лемоцита (пунктиром показана плоскость поперечного разреза);
7 - контакт двух лемоцитив.
Безмиелиновые нервные волокна тонкие, их диаметр составляет 1-4 мкм. Они типичны для автономной (вегетативной) нервной системы. Характерной их структурным признаком является то, что отростки нейронов (осевые цилиндры) прогибает плазматической мембраной олигодендроцита (нейролемоцита) и погружаются в него, образуется глиальные муфта вокруг нервного отростка (рис. 1). В одну клетку Шванна может быть погружено много нервных волокон. Однако некоторые волокна не покрыты со всех сторон олигодендроцитамы. Группа безмиелинових нервных волокон, которая связана с одним нейролемоцитом, покрытая ендоневрием, образованным базальной мембраной нейролемоцита и тонкой сеточкой из коллагеновых и ретикулярных микрофибрилл.
Миелиновые нервные волокна имеют сложное строение потому нейролемоцит (клетка Шванна) спирально накручивается на осевой цилиндр (аксон) нервной клетки. При этом цитоплазма и ядро нейролемоцита оттеснены на периферию в поверхностные отделы оболочки (рис. 2). Это толстые волокна диаметром от 1 до 20 мкм. Они являются составной частью центральной и периферической нервной системы. Каждый нейролемоцит охватывает только часть осевого цилиндра длиной примерно 1 мм, образуя межузловых сегмент нервного волокна. Миелин - это многократно закрученный двойной слой цитоплазматической мембраны нейролемоцита. Толстая и плотная миелиновая оболочка, богатая липидами, изолирует нервное волокно и предотвращает "утечки" электрического тока (нервного импульса) с аксолемы. Внешняя оболочка осевого цилиндра образована цитолемою нейролемоцита, его базальной мембраной и тонкой сеточкой из ретикулярных и коллагеновых фибрилл. В темной миелиновых оболочке (при окраске препаратов осмиево кислотой) расположены узкие светлые косые линии - насечки миелина. Эти структуры связаны с процессом формирования миелиновой оболочки. Двойная складка плазмолемы нейролемоцита, что концентрически расположена вокруг осевого цилиндра, называется мезаксоном. Каждый завиток мезаксона имеет толщиной 8-12 нм. Насечки миелина соответствуют местам, где завитки мезаксона раздвинуты цитоплазмой лемоцитов. На границе между двумя соседними нейролемоцитамы образуется сужение нервного волокна - узловая перетяжка нервного волокна (перетяжка Ранвье), шириной в среднем 0,5 мкм, где миелиновая оболочка отсутствует (рис.3 ). Здесь аксолема контактирует с отростками, переплетающихся между собой, нейролемоцитамы и их базальной мембраной. В этом месте сконцентрировано большинство натриевых каналов (3000-5000 на 1 мкм), а в цитолеми, покрытой миелина, они практически отсутствуют. Междоузлия сегменты покрыты миелина, поэтому время проведения по ним нервного импульса приближается к нулю.
В аксолеми на уровнях перетяжек Ранвье генерируется нервный импульс, стремительно проводится в соседнюю перетяжки, в которой возбуждается следующий потенциал действия. Такой способ проведения импульса называется сальтаторним (скачкообразным). В миелиновых нервных волокнах скорость проведения нервного импульса высокая - от 5 до 120 м / с. Безмиелинови нервные волокна проводят нервный импульс со скоростью всего 1-2 м / с, потому что волна деполяризации движется по всей плазмолеми, не прерываясь.
Между нейронами и глиоцитамы является межклеточные щели шириной 15-20 нм. Они заполнены основной межклеточной веществом из мукополисахаридов и других соединений, обеспечивающих диффузию кислорода и питательных веществ. Все межклеточные щели соединяются между собой и образуют межклеточное (интерстициальное) пространство, на который приходится 17-20% общего объема мозга.
В зависимости от функции выделяют три основных типа нейронов: афферентные, ассоциативные и эфферентные. Афферентные нейроны (от латинского affevens - тот, что приносит) по функции чувствительны (рецепторными) клетками, они проводят нервные импульсы от органов и тканей в мозг. Тела таких нейронов сосредоточены в нервных узлах (ганглиях), расположенные вне центральной нервной системой. Как правило, это биполярные (псевдоуниполярные) нейроны. Долгое дендрит такой нервной клетки направляется на периферию и заканчивается соответствующим рецептором, который способен трансформировать энергию внешнего раздражителя в нервный импульс. Второй отросток (аксон) идет в головной или спинной мозг, принося соответствующую информацию.
Рецепторы (receptores) в зависимости от их локализации разделяют на: екстерорецепторы, воспринимающие внешние раздражения, - они расположены в наружных покровах тела (коже и слизистых оболочках, в органах чувств); интерорецепторы, которые разделяют на те, которые реагируют на изменение химического состава внутренней среды (хеморецепторы), давления в тканях и органах (барорецепторов, механорецепторов); проприорецепторы, расположенных в мышц, сухожилий, связках, фасциях, костях, суставных капсулах и т.д. и реагируют на структурно-функциональные изменения в опорно-двигательном аппарате .
В зависимости от характера восприятия раздражения различают: терморецепторы, механорецепторов, барорецепторов, хеморецепторы и ноцирецепторы. Терморецепторы воспринимают изменение температуры. Механорецепторов реагируют на различные виды механических воздействий (прикосновение к коже, ее сжатия и др.), ноцирецепторы воспринимают болевые раздражения. Барорецепторов воспринимают изменения давления, а хеморецепторы реагируют на воздействие химических факторов. Среди нервных окончаний различают свободные, состоящие только из разветвлений осевого цилиндра, и несвободные, или конечные нервные тельца, содержащие окончания нервного волокна и клетки глии.
Свободные нервные окончания (termindtio nervi liberae) расположены в коже. Подходя к эпидермиса, нервное волокно теряет миелин и проникает через базальную мембрану в эпителиальный слой. При этом базальные мембраны эпителия и нейролемоцитив переходят друг в друга. Нервные волокна разветвляются между эпителиоцитами, доходя до зернистого слоя; их веточки диаметром менее 0,2 мкм колбоподибно расширяются на концах. Аналогичные конечные нервные окончания есть в эпителии слизистых оболочек и роговице глаза. Конечные свободные нервные окончания воспринимают боль, тепло и холод.
В эпидермисе свободные нервные окончания подходят к изменившимся эпителиоцитов (клеток Меркеля), которые имеют много пальцеподибних выростов. В цитоплазме этих клеток много електроннощильних мембранных гранул диаметром приблизительно 100 нм. Нервное окончание расширяется и образует из клеток Меркеля синапсоподибний контакт. Эти окончания является механорецепторов и воспринимают давление.
Барорецепторов - это разветвленные свободные нервные окончания, расположенные в адвентицийному слое стенки крупных артерий грудной полости и шеи. Важнейшие рецепторы залегают в стенках дуги аорты и сонной пазухи. При растяжении стенки артерии под влиянием артериального давления, что меняется, они возбуждаются, а закодированная информация о величине давления крови передается в центральную нервную систему. Таким образом, барорецепторов обеспечивают регуляцию артериального давления по принципу обратной связи.
Несвободные нервные окончания могут быть капсулированных, т.е. окружены соединительнотканной капсулой, и некапсульованимы, которые не имеют капсулы. Некапсульовани нервные окончания (corpusculum nervosum попcapsulatum) расположены в соединительной ткани. К ним относятся также окончание в волосяных фолликулах. Капсулированных нервными окончаниями (corpusculum nervosum capsulatum) является осязательные тельца (осязательные тельца Мейснера), пластинчатые тельца (тельца Фатер-Пачино), цибулиноподибни тельца (тельца Гольджи-Маццони). Все эти нервные окончания есть механорецепторов. В группу капсулированных нервных окончаний принадлежат колбочки Краузе, которые являются терморецепторы.
Каждый волосяной фолликул окружен расширенными несвободными нервными окончаниями, которые расположены снаружи от соединительнотканных элементов влагалища корня волоса. Базальная мембрана нервных окончаний переходит в базальную мембрану внешней эпителиальной влагалища. Уплощенное нервное окончание залегает между двумя клетками Шванна. При движении во-лосины возникает нервный импульс и передается в центральную нервную систему.
Пластинчатые тельца (тельца Фатер-Пачино) - крупнейшие из всех капсулированных нервных окончаний. Они имеют овальную форму, длину до 2 мм и толщину 0,5 мм. Расположенные в соединительной ткани внутренних органов и в подкожной основе (рис. 4,5). Тельце извне покрыто соединительнотканной капсулой, имеющей пластинчатые строение. Во соединительно тканевой оболочкой размещена внешняя луковица, состоящая из 10-60 концентрических пластинок, образованных уплощенным периневральным эпителиоидных клетками, соединенными между собой десмосомамы и замыкающими зонами.
Пространства между пластинками заполнены жидкостью, коллагеновыми микрофибрилл и редкими кровеносными капиллярами. Войдя в тельце, нервное волокно теряет миелиновую оболочку, но внутри тельца оно окружено клетками Шванна, формирующие внутреннюю луковицу. Нервное волокно, колбоподибно заканчивается, содержит много маленьких сферических митохондрий и светлых синаптических пузырьков. При сжатии тельца в нервном окончании возникает нервный импульс.
Осязательные тельца (осязательные тельца Мейснера) длина 50-160 мкм, шириной примерно бОмкм, имеют овальную или цилиндрическую форму, их много в сосочковыми слое кожи пальцев рук и стопы. Тельце образованное многочисленными удлиненными или грушевидными клетками Шванна, лежащие одна на одной. Нервное волокно, входя в тельце, теряет миелин и располагается между цитоплазматическими отростками. Каждое волокно заканчивается колбоподибним расширением. Периневрий переходит в капсулу, которая образована несколькими слоями эпителиоидных периневральным клеток. Сжатие этих клеток передается осязательную тельцу, вызывая деформацию его клеток, что приводит к возникновению импульса в нервных волокнах. То есть, осязательные тельца Мейснера является механорецепторов тактильной чувствительности.
Цибулиноподибни тельца (колбочки Краузе) расположены в коже, коньюнктивы глаза, слизистой оболочке полости рта. Сферические тельца окружены тонкой соединительнотканной капсулой, содержащей много коллагеновых волокон и фибробластов. Войдя в капсулу, нервное волокно теряет миелиновую оболочку и разветвляется в центре колбы на многочисленные веточки. Колбочки Краузе воспринимают холод, возможно, они есть и механорецепторов.
Проприорецепторы реагируют на напряжение мышц, сухожилий и суставных капсул, возникающие при движениях и изменении положения тела в пространстве. Это нервно-мышечные и нервно-сухожилий веретена, расположенные в брюшке мышц или в их сухожилия.
Большое (длиной 3-5 мм и толщиной до 0,5 мм) нервно-мышечное веретено окружено плотной соединительнотканной внутренней капсулой и состоит из 10-12 поперечно исполосованная мышечных волокон, а также нервных волокон. Наружная капсула веретена формируется из поперечно исполосованная мышечных волокон. Продольно расположенные веретена растягиваются вместе с мышцей и передают информацию о длине и степень растяжения мышцы.
Вставные (ассоциативные или кондукторни) нейроны передают нервный импульс от чувствительного (центростремительной) нейрона к двигательного или секреторного (центробежного) нейрона.
Тела эфферентных (эффекторных; двигательных; секреторных) нейронов (от латинского effevens - тот, который выносит) расположены в центральной нервной системе, симпатичных и парасимпатических нервных узлах. Аксоны эфферентных нейронов проводят нервные импульсы в рабочие органы (мышц, желез), а также ко всем органам и тканям, регулируя таким образом обмен веществ. Выделяют также двигательные и секреторные окончания аксонов эфферентных нейронов. Аксоны двигательных нейронов разветвляются, каждая такая ветка иннервируют определенное количество мышечных волокон. Двигательные (моторные) нервные окончания располагаются на мышечных волокнах. Окончание одного двигательного нейрона и поперечно исполосованная мышечное волокно, что им иннервуеться, образуют двигательную единицу - мион.
Каждое мышечное волокно иннервируют веточка аксона двигательного нейрона, которая заканчивается на поверхности волокна, образуя нервно-мышечное окончание, аборухову конечную пластинку - моторную бляшку (рис. 6). Веточки аксонов, контактирующие с мышечным волок-ном, уже не имеют миелиновой оболочки. Каждое такое нервное окончание прогибает сарколему мышечного волокна и образует нервно-мышечный синапс. Нервно-мышечный синапс состоит из пресинаптических мембраны (аксолема) и постсинаптические мембраны (сарколема мышечного волокна), разделенных синаптической щели (рис. 7).
Нервный импульс, достигающий нервно-мышечного синапса, вызывает выход в синаптическую щель ацетилхолина, который связывается со специфическими рецепторами мембраны мышечного волокна, что приводит к открытию натриевого и калиевого каналов. Это вызывает местную деполяризацию постсинаптические мембраны, а электрический потенциал запускает механизм сокращения мышечного волокна.
Нервные окончания в гладкой мышечной ткани имеют более простую строение, образуя на поверхности миоциты характерные расширения, в которых есть синаптическую пузырьки с норадреналин и дофамин. Большинство окончаний аксонов контактируют с базальной мембраной гладкого миоциты, лишь незначительное количество нервных окончаний пронизывает базальную мембрану Тогда аксолема отделена от плазмолемы миоциты щелью шириной около 10 нм.
Рис. 1. Строение безмиелинового нервного волокна
I - продольный разрез;
II - поперечный разрез.
1 - осевые цилиндры;
2 - аксолема;
3 - мезаксон;
4 - клеточная оболочка лемоцита (нейролемоцита; клетки Шванна);
5 - цитоплазма лемоцита;
6 - ядро лемоцита (пунктиром показана плоскость поперечного разреза);
7 - контакт двух лемоцитив.
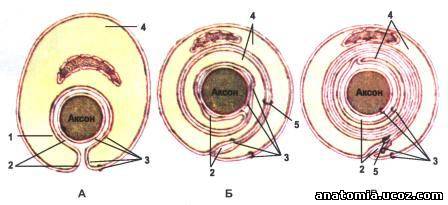
Рис. 2. Последовательные стадии (А, Б, В) формирование миелинового нервного волокна.
1 - контакт аксолемы и клеточной оболочки лемоцита (нейролемоцита; клетки Шванна); 2 - межклеточная щель; 3 - аксолема и клеточная оболочка лемоцита; 4 - цитоплазма лемоцита; 5 - мезаксон.
1 - контакт аксолемы и клеточной оболочки лемоцита (нейролемоцита; клетки Шванна); 2 - межклеточная щель; 3 - аксолема и клеточная оболочка лемоцита; 4 - цитоплазма лемоцита; 5 - мезаксон.
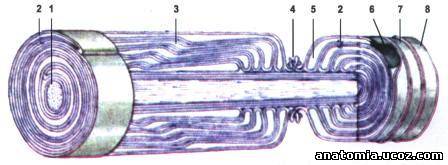
Рис. 3. Строение миелинового нервного волокна и перетяжки Ранвье.
1 - осевой цилиндр; 2 - мезаксон; 3 - насечки миелина; 4 - перетяжки Ранвье (узловая перетяжка нервного волокна); 5 - цитоплазма лемоцита; 6 - ядро лемоцита; 7 - нейролема; 8 - ендоневрий.
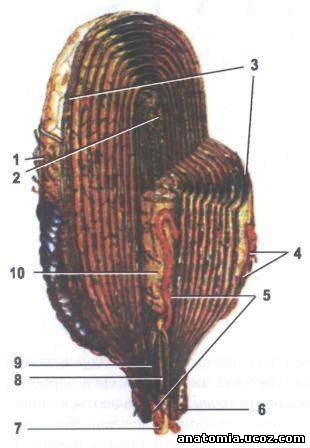
Рис. 4. Тельце Фатер-Пачино - продольный разрез:
1 - соединительная капсула;
2 - окончание нервного волокна;
3 - пластинки внешней луковицы;
4 - коллагеновые волокна;
5 - гемокапиляры;
6 - периневрий;
7 - миелиновая нервное волокно;
8 - нервное волокно (дендрит);
9 - миелин;
10 - внутренняя луковица.
1 - соединительная капсула;
2 - окончание нервного волокна;
3 - пластинки внешней луковицы;
4 - коллагеновые волокна;
5 - гемокапиляры;
6 - периневрий;
7 - миелиновая нервное волокно;
8 - нервное волокно (дендрит);
9 - миелин;
10 - внутренняя луковица.
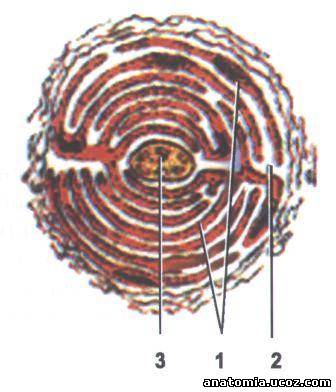
Рис. 5. Тельце Фатер-Пачино - поперечный разрез:
1 - пластинка внутренней луковицы;
2 - радиальная щель;
3 - нервное волокно.
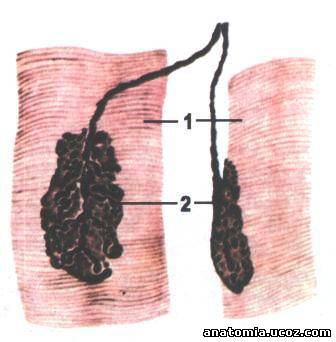
Рис. 6. Нервно-мышечное окончание (двигательная конечная пластинка).
1 - мышечное волокно; 2 - моторная бляшка.
1 - мышечное волокно; 2 - моторная бляшка.
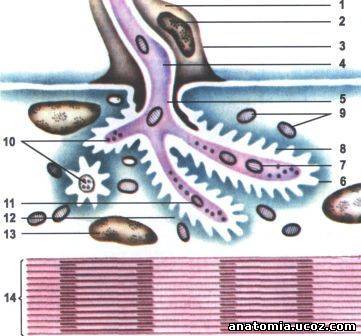
Рис. 7. Строение нервно-мышечного синапса
1 - нейролемоцит;
2 - ядро нейролемоцита;
3 - нейролема (клеточная мембрана нейролемоцита);
4 - аксоплазма;
5 - аксолема;
6 - митохондрии мышечного волокна;
7 - синаптическая щель;
8 - митохондрии нервного окончания;
9 - сарколема (постсинаптические мембрана);
10 - синаптическую пузырьки;
11 - аксолема (пресинаптических мембран);
12 - сарколема;
13 - ядро мышечного волокна;
14 - миофибриллы.